畜牧设备有限公司")
影响奶牛乳蛋白质含量的因素及营养调控技术研究
- 2016-10-12 16:49:00
- administrator 原创
- 4036
1 乳中蛋白质的合成
1.1 乳腺中合成的蛋白质
乳腺是合成乳汁的主要场所。乳中90%以上的蛋白质是在乳腺中由氨基酸从头合成的。乳蛋白合成所需要的蛋白质占蛋白质总需要的70%。酪蛋白是乳中主要的蛋白质之一,约占乳中粗蛋白总量的80%(DePeters等,1992),主要由乳腺从血液中摄取的游离的氨基酸以及小分子寡肽合成(Backwell等,1996),其组成成分(α-酪蛋白、β-酪蛋白、κ-酪蛋白、γ-酪蛋白)对乳品品质有很大的影响。另外,乳腺吸收的氨基酸除了用以合成酪蛋白外,还用来合成β-乳球蛋白和α-乳清蛋白(Clark等,1978)。与酪蛋白不同,有些乳清蛋白则完全由乳腺组织吸收。乳腺组织从血液中摄取的氨基酸量与合成乳蛋白的氨基酸量并不一致,有的氨基酸(Lys、Arg、Ile、Leu 和Val)摄入量高于输出量,而有的氨基酸(Met、Phe、His和 Thr)摄入量低于输出量(Bequette,1997;Guinard,1994;Mabjeesh,2002)。这可能说明,除了血液中游离的氨基酸参与乳腺组织氨基酸的供给外,肽和蛋白质也参与乳腺组织氨基酸的供给,用来合成乳中蛋白质,这部分肽和蛋白质刚好可以弥补乳腺组织摄入游离氨基酸的不足(Bequette,1999;Mabjeesh,2000)。而乳腺组织摄入过量的氨基酸除直接用于合成乳蛋白外,仍有一部分氨基酸参与合成其它功能物质(合成细胞的结构蛋白或酶类;合成非必需氨基酸、多胺、CO2和ATP)和支链氨基酸(Bequette ,1998)。
乳腺组织蛋白质的合成与乳腺对氨基酸的摄取有关,而控制乳腺从血液中摄取游离氨基酸的主要因素是:①乳腺氨基酸转运系统的活力;②乳腺摄入的血浆游离氨基酸量占血浆总氨基酸量的百分比(Km);③血浆游离氨基酸的净摄入量。但多数研究认为,乳腺对氨基酸的摄取是相对保守的,不同的氨基酸转运系统活力不同,Km值也不同。正常情况下,它们会维持在一个特定的范围内;当过量添加游离氨基酸时,它们会通过下调而维持血浆游离氨基酸净摄入量的稳定。
1.2 乳中的血液蛋白质
牛乳中5%~10%的乳蛋白质来自血液,一种是血清清蛋白,存在于牛乳乳清中,与血液中的血清蛋白完全相同;另一种是免疫球蛋白,初乳中的免疫球蛋白大部分来自血液。就母羊而言,每升初乳中的免疫球蛋白高达100多克,而常乳中的免疫球蛋白水平低于初乳。另外,不同种别的动物,初乳中的免疫球蛋白的种类不同,如在牛的初乳中,主要是IgG、IgM和IgA 3种成分,分别占免疫球蛋白总量的85%~90%、10%和5%左右。
2 影响牛奶中蛋白质含量和产量的因素
2.1 非营养因素
2.1.1 品种
相对于乳脂率而言,乳蛋白含量受遗传影响较大。奶牛品种是影响乳蛋白含量的主要因素,品种不同,乳中蛋白质含量和产量也不同。中国荷斯坦奶牛平均乳蛋白含量最低(3.1%);娟姗牛的乳蛋白含量最高(4.0%以上);爱尔夏牛、黑白花牛和更赛牛等乳蛋白含量适中。一般而言,产奶量越高,乳蛋白含量就越低,乳品品质就越差。同一品种内的不同品系间的乳蛋白含量也存在较大的差异。
2.1.2 泌乳阶段
同一泌乳期内,泌乳各个月的乳蛋白含量变化很大。产犊后的11d乳成分变化最大,尤其是乳蛋白率和总固体率,其次是乳脂率,而乳糖、灰分则相对稳定。牛乳中蛋白质在泌乳初期和后期较高,在产犊后的5~10周,则乳中蛋白质含量,酪蛋白及非蛋白氮(NPN)含量快速下降,随后在泌乳末期逐渐上升(Ng-Kwai Hang,1984、1985)。泌乳阶段对乳中蛋白质产量的影响小于对脂肪产量的影响。一般来说,产奶量高时乳脂率低,产奶量低时乳脂率高,乳脂率与乳蛋白率的升降有相似的趋势。可用NRC(1978)公式由乳脂率计算乳蛋白率:乳蛋白率(%)=1.9+0.4×乳脂率(%)。如果饲料中由于纤维量不够或粗饲料被粉碎而导致乳脂率下降,则不可估算乳蛋白率。
2.1.3 环境温度
环境温度对乳中蛋白质含量有一定的影响,环境温度过高则乳中蛋白质含量下降(Fegan,1979)。Bruhn等(1977)对4个不同品种的奶牛试验的研究结果表明,夏季乳中蛋白质含量最低而冬季最高,但牛奶中乳蛋白质含量的变化小于乳脂的变化。夏季和冬季乳蛋白质含量变化范围在0.2%~0.3%,环境温度对乳蛋白含量的影响是否与高温或昼夜时间的长短有关还不太清楚,有待进一步去研究。
2.1.4 胎次
奶牛的胎次对乳蛋白含量的影响不太明显。随着胎次的增加,乳蛋白质含量有下降趋势,但下降幅度很小。一般来说,第一胎时乳蛋白含量有下降趋势,第二胎以后则乳蛋白含量的变化基本稳定。
2.1.5 疾病
临床的乳房炎和其它炎症都会使奶中酪蛋白含量下降,乳清蛋白质含量却上升(Fegan,1979;Munro,1984),这是由于牛奶中体细胞数量增多,纤维蛋白溶解酶、碱性乳蛋白酶的活性与正常牛奶相比增高的缘故。患乳房炎时,血浆蛋白质进入牛奶中的量增加,牛奶的pH值升高,碱性增强,这可能是纤维蛋白质溶解酶含量或活性升高而导致酪蛋白的分解增加。
2.2 营养因素
2.2.1 能量
最近,越来越多的研究表明,乳蛋白含量与能量摄入量之间存在一定的相关性。Emery(1978)就指出,当由谷物或粗饲料提供的净能增加4.18MJ时,乳蛋白含量增加0.015单位,能量的摄入量与乳蛋白含量呈正相关(r=0.42)。Sporndly(1989)的研究同样表明,代谢能(ME)的摄入量与乳蛋白含量或产量之间存在正相关(r=0.42和0.31),日粮每提供1MJ 的代谢能则乳蛋白含量增加0.03单位,相当于每提供4.18MJ的净能则乳蛋白提高0.02单位。这说明乳蛋白含量主要受日粮能量水平的影响。日粮能量和碳水化合物对乳脂和乳蛋白的影响主要取决于瘤胃内发酵生成的乙酸盐和丙酸盐的比例,而这种比例是由日粮能量和碳水化合物水平及发酵类型决定的。能量不足时,一方面降低了瘤胃微生物蛋白质合成量,使进入小肠内的瘤胃微生物蛋白减少,乳腺中乳蛋白的合成减弱;另一方面导致合成乳蛋白的氨基酸被当作能量利用,从而使乳蛋白浓度下降。反之,增加日粮中的能量和碳水化合物,用于供能的氨基酸减少,微生物蛋白质合成增多,同样使瘤胃中丙酸盐比例提高,刺激胰岛素分泌,增加乳腺对氨基酸的吸收,从而提高乳蛋白率。而影响日粮能量水平的决定因素是日粮中精料的含量和牧草的青贮品质。因此,通过提高精料水平或改进青贮品质可增加奶牛的能量摄入量,从而提高乳蛋白含量和产量。
2.2.2 脂肪
日粮的能量是由需要量和采食量来决定的。如果不减少纤维的含量,产奶量大于35kg/d时,就要补饲脂肪。Palmquist(1993)建议日粮内脂肪的上限应相当于乳中的脂肪含量。日粮脂肪水平一般控制在干物质的7%~9%水平以下。日粮脂肪的1/3来自基础日粮组分(谷物、牧草);1/3来自油类籽实(棉籽、大豆等);其余由惰性脂肪提供,具体要求见表1(卢德勋,2001)。如果不是用惰性脂肪,日粮含脂肪4%~5%时就会导致瘤胃发酵改变,如降低纤维素的消化率和乙、丙酸盐比例(William Chalupa,1984)。试验证实,奶牛日粮添加脂肪后虽然增加了产奶量,但常会导致乳蛋白率下降(Emery,1978),而酪蛋白是乳中含氮组分中下降最明显的成分(Depeters等,1992)。杨金波等(2002)报道,根据泌乳奶牛产奶量的差异,在其精料中添加适量的过瘤胃脂肪,乳蛋白的含量下降了4.51%。Sporndly(1989)研究发现,日粮中的粗脂肪含量与乳蛋白产量呈正相关(r=0.31),而与乳蛋白含量呈负相关(r=-0.24)。Wu和Huber(1994)分析了大量添加脂肪试验的研究结果,发现随着日粮脂肪添加量的增加,乳蛋白产量不变或提高,而乳蛋白含量却比对照日粮(脂肪含量为3%)有所下降。另有学者的分析结果是,当日粮脂肪酸从2.1%增加到10.7%,每增加100g脂肪则乳蛋白含量下降0.03个百分点(Pamlquist, 1993)。

日粮添加脂肪导致乳蛋白率下降的解释很多。一种观点是这种下降与葡萄糖代谢有关,即添加脂肪后日粮中谷物比例降低,导致葡萄糖前体减少,会增加氨基酸的葡萄糖异生作用,从而降低乳蛋白合成量。Cant等(1991)对初产母牛饲喂脂肪含量分别为2%和5%的两种日粮,日粮蛋白质含量为15.4%,结果乳蛋白率分别为3.9%和2.96%;经皱胃灌注酪蛋白后,5%脂肪日粮的乳蛋白率提高到3.04%。因此认为,灌注酪蛋白后除可以增加日粮蛋白质量,改善氨基酸的平衡外,还可以克服由于补饲脂肪后奶牛日粮采食量下降,大量氨基酸的葡萄糖异生作用造成的乳蛋白率下降。另一种观点是应用保护或非保护性脂肪降低乳蛋白率可能是由于日粮含碳水化合物过少,使微生物蛋白质的合成量减少。
3 乳中蛋白质调控的营养途径 3.1 日粮碳水化合物 3.3 控制蛋白质水平 3.4 瘤胃保护性氨基酸
2.2.3 蛋白质
蛋白质是乳蛋白合成的主要原料,也是泌乳奶牛日粮的主要限制性营养成分之一。蛋白质在瘤胃中被瘤胃微生物降解并合成微生物蛋白,与瘤胃未降解蛋白、内源性蛋白随瘤胃食糜进入小肠,组成小肠代谢蛋白,在小肠中被消化吸收,并在乳腺中合成乳蛋白。但是,瘤胃微生物蛋白合成效率不高,并且氨基酸组成不平衡,从而影响了乳蛋白的合成。因此,日粮中蛋白质缺乏导致乳蛋白浓度下降,而补充蛋白质可显著提高乳蛋白浓度。Emery(1978)发现,日粮蛋白质每增加一个百分点,牛乳中蛋白质的水平就增加0.02个百分点。但前提是在奶牛日粮蛋白严重不足的条件下,增加日粮蛋白质水平可提高奶牛的消化率和采食量,所以产奶量也随之上升,导致乳蛋白含量的增加。因此,可以认为产奶量和乳蛋白含量的增加,是饲料蛋白质直接效应和能量间接效应的双重结果。另一方面,日粮中补充过瘤胃蛋白质饲料可以改善真胃和小肠蛋白质营养,并维持较高的乳蛋白质产量。奶牛饲料中的瘤胃非降解蛋白质(过瘤胃蛋白)和微生物蛋白是奶牛小肠蛋白质的主要来源,而且日粮中到达真胃和小肠的非降解蛋白质或氨基酸不足是影响高产奶牛乳蛋白含量的主要因素。殷光骐等(1999)在奶牛日粮中增加非降解蛋白饲料的试验表明,试验组比对照组的乳蛋白率略有提高(P>0.05),且很少受其它饲料及天气的影响,含量稳定。同样,在常规日粮条件下通过真胃或十二指肠灌注酪蛋白也能明显提高产奶量和乳蛋白产量(见表2),其原因除增加了日粮中蛋白量外还改善了氨基酸的平衡。因此,只有保持微生物蛋白和过瘤胃蛋白的平衡,才能使乳腺获得合成乳蛋白的理想蛋白质。
对于奶牛的营养管理来说,在奶牛饲养方式上多采取粗料自由采食,精料则根据奶牛的生产水平来确定适宜的添加量,以提高乳蛋白为目标来设计营养调控型饲料产品和平衡日粮。比如适宜的日粮精粗比(60:40);适宜的日粮淀粉水平和非纤维碳水化合物(NFC,40%~45%);日粮粗蛋白水平;日粮过瘤胃蛋白(33%~40%)等。但以哪些指标来设计营养调控型饲料产品和平衡日粮是目前研究者和生产者所关心的问题。多数研究表明,在正常情况下,乳蛋白含量反映了泌乳牛的能量平衡状况(与过瘤胃蛋白质的摄入量也有一定关系),而乳尿素氮含量反映日粮蛋白质摄入的平衡状况(Hof等,1997;Hwang等,2001)(见表3)。因此,在生产实践中,可以根据乳蛋白和乳尿素氮的含量来衡量奶牛日粮中能量和蛋白质的平衡状况,进而及时调整日粮,以提高乳蛋白率。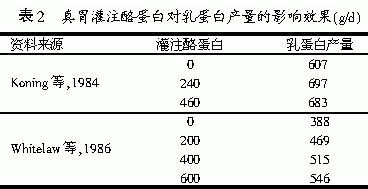
碳水化合物分为结构碳水化合物和非结构碳水化合物两大类,通常占饲粮比例的60%~70%。非结构碳水化合物(NSC)存在于植物细胞内部,主要是细胞内容物中的淀粉和糖,通常比结构碳水化合物更容易被消化,在泌乳奶牛饲粮中用来替代部分中性洗涤纤维(NDF),以满足其对能量的需要。因此,在实际生产中,为了给高产奶牛或泌乳高峰期奶牛提供所需的能量,常常减少配合日粮粗纤维的比例,增加非纤维性碳水化合物(NFC)的含量。NFC包括淀粉、糖和果胶,通常用下面公式来计算:NFC=100%-(NDF%+CP%+Fat%+Ash%)(NDF表示中性洗涤纤维;CP表示粗蛋白;Fat表示粗脂肪;Ash表示灰分)(孟庆翔,2002)。
改变饲粮NFC含量能够影响瘤胃的发酵模式、纤维在整个消化道的消化和乳成分(Sievent和Shaver,1993;Sutton和Bines,1987)。如饲喂适量的NFC可以促进丙酸的合成,乙酸和丙酸的比例相应下降,乳脂率下降,乳蛋白率升高。Hoover和Stokes(1991)将Nocek和Russell(1988)得到的数据进行了回归分析,发现饲粮中NFC比例在大于45%~50%和小于25%~30%时,产奶量下降。Minor等(1998)研究发现,当NFC在饲粮干物质中的比例由41.7%增加到46.5%时,乳蛋白的含量和产量都有增加。因此,为满足奶牛生产的需要,奶牛饲粮中应含有适宜的NFC比例,否则会导致瘤胃酸中毒和其它代谢疾病的发生。一般来说,饲粮中NSC的最大比例为饲粮干物质(DM)的30%~40%(Nocke,1997)。对NFC来说,在奶牛饲粮中可以接受的比例大约比NSC高2~3个百分点。
3.2 调节日粮精粗比
多数研究与饲养实践表明,奶牛能量的摄入量可通过改变日粮的精粗比来实现。通过增加精料水平提高精粗比,来增加能量摄入量,而增加的能量可提高乳蛋白的含量和产量(Macleod等,1983)。因此,在奶牛饲养方式上采取粗料自由采食,通过增加精料水平提高精粗比,从而提高乳蛋白含量和产量。研究表明,对以青贮为基础日粮的母牛,每天补充3.0、6.0、9.0kg精料,乳蛋白率可从3.08%提高到3.31%。Bartsch等(1979)的试验结果同样证实了这一结论(见表4)。尽管如此,提高日粮精粗比在提高乳蛋白含量和产量的同时,却对乳中的其它成分有不良影响,如高浓度精料的日粮降低乳脂率(王加启,1994;李坚,2001;张廷利,1997;Ireland perry和stallings,1993;Grummer,1987;Taquette,1986Zimmerman,1991 Khorassami,2001 Poore,1993),主要是由于高浓度的精料日粮会使瘤胃发酵类型改变,由高乙酸型转变为高丙酸型(王加启,1993)。因此,在实际生产中,应根据生产需要来调整适宜的精粗比。一般精粗比为60:40,否则会造成酸中毒,进而降低采食量。由于奶牛乳蛋白的适宜日粮精粗比受日粮中粗料和精料的组成及品质的影响,得到既不影响乳蛋白含量和产量,又不影响乳脂率的适宜精粗比,是当今研究者和生产者所面临的挑战。根据日粮内粗料的组成和品质来调整适宜的精粗比,具体要求见表5。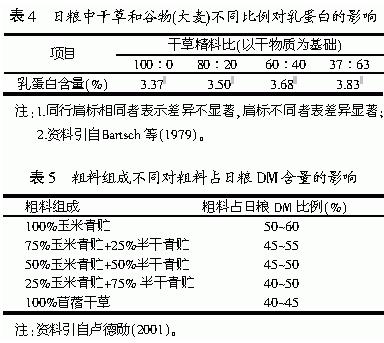
目前调控乳蛋白的方法主要是控制日粮蛋白质水平和瘤胃非降解蛋白(RUP)水平。在饲养实践中,由于不可能对每份饲料的营养成分进行实测,因此往往采取在理论蛋白质需要量的基础上再增加10%~15%的做法,以保证奶牛的营养需要来改善乳蛋白的合成。但结果不十分理想,由于大部分日粮蛋白在瘤胃中被降解浪费掉,因而在日粮蛋白满足奶牛营养需要的条件下,再提高日粮蛋白水平,并不是调控乳蛋白的有效途径。随后研究者又发现,乳蛋白的合成受小肠蛋白质供给量或多种氨基酸的限制,而小肠蛋白质的供给受日粮中过瘤胃蛋白和微生物蛋白的合成和流失的影响,只有保持微生物蛋白和过瘤胃蛋白的平衡,才能使乳腺获得合成乳蛋白的理想蛋白质,这样才能使乳蛋白合成达到最优化。因此,饲料中含有适量的降解率低的蛋白饲料是调控乳蛋白的有效途径之一。此外,在使用的时候还应该注意它们的添加量,日粮中瘤胃非降解蛋白含量过高、过低都不好。Rodriguez等(1997)指出,奶牛日粮中的瘤胃非降解蛋白一般不能超过40%,高水平的RUP(41%)可降低乳蛋白率。总之,保持瘤胃降解蛋白和未降解蛋白的平衡是调控乳蛋白的有利保证。奶牛对日粮蛋白质部分的营养要求如表6所示。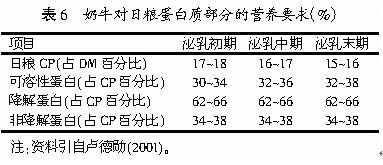
通过添加低降解蛋白质饲料来提高进入小肠的可利用蛋白量,满足奶牛乳蛋白合成的需要,效果并不十分理想,造成这种现象的主要原因可能是添加的低降解蛋白质饲料所含的氨基酸与机体所缺的氨基酸不能完全匹配。因为乳蛋白的合成与供给乳腺氨基酸的组成有关,于是人们开始在奶牛日粮中补充限制性氨基酸,保证乳腺合成乳蛋白所需要的理想氨基酸组成,以便使乳腺蛋白质合成达到最优化。
目前绝大多数对泌乳奶牛的限制性氨基酸的鉴别工作表明,赖氨酸和蛋氨酸是代谢蛋白质(MP)第一限制性氨基酸,对于蛋氨酸和赖氨酸以外的其它限制性氨基酸的确定工作还很有限。尽管如此,对于不同泌乳阶段与饲粮组分的奶牛来说,其限制性氨基酸可能有所差异。
Boisen等(2000)研究了不同日粮的第一限制性氨基酸,对菜籽粕来说,亮氨酸是第一限制性氨基酸;对谷类饲料赖氨酸是第一限制性氨基酸;豆粕日粮中蛋氨酸是第一限制性氨基酸;由大麦、玉米、菜籽粕和干草配制的混合日粮,赖氨酸是第一限制性氨基酸。因此,在奶牛日粮中添加限制性氨基酸的时候,应根据其泌乳阶段和日粮组成添加相应的限制性氨基酸才能达到预期的目的。目前,使用最多的是瘤胃保护性蛋氨酸和赖氨酸这两种氨基酸。总的来说,奶牛日粮中添加瘤胃保护性氨基酸均不同程度地提高乳蛋白含量和产量,但效果也不尽一致。可能的原因是:①日粮组成不同;②瘤胃保护性氨基酸的降解率(保护形式)不同;③瘤胃保护性氨基酸的添加量不同。
3.5 维生素
B族维生素在蛋白质合成、氧化分解及其它代谢过程中起着重要作用,饲料中它们的含量对乳蛋白具有一定的影响。尽管奶牛瘤胃微生物能够合成B族维生素,但对于高产的奶牛而言,瘤胃微生物合成的维生素是不能满足需要的。因此,在奶牛日粮中添加B族维生素,可在一定程度上改善奶牛产奶性能,提高乳品品质。Grummer等(1987)认为,奶牛日粮中补充叶酸,可以使血清中叶酸浓度升高,导致瘤胃内丙酸产量增加,进而提高乳蛋白质产量 。而目前在奶牛生产中应用最多的是烟酸。多数研究认为,奶牛日粮中补充烟酸,可提高乳蛋白的含量(Horner等,1986)。另外,研究者和奶牛生产者通过在奶牛高脂日粮中补充烟酸,来有效地缓解或提高因补充脂肪而引起的乳蛋白含量降低问题,即使饲喂普通(低脂)日粮,补充烟酸也可提高乳蛋白的含量。实际生产中建议每头奶牛每日补饲烟酸6g(每千克干物质200~400mg),且补饲期应从产犊前1周开始直到配种。关于烟酸提高乳蛋白含量的机制大致有以下几种解释:①提高微生物蛋白质合成效率或增加微生物生长所需能量,因而增加了十二指肠微生物氮量,提高了小肠氨基酸的利用率(Erickson等,1992);②提高了瘤胃中丙酸盐浓度,使乙酸和丁酸浓度降低(Flachowsky等,1988;Riddle等,1981);③减少了奶牛对高精料饲粮的应激。牛采食高精料会引起应激反应, 导致采食量下降, 补饲烟酸有益于改善高精料的不良作用。不过,确切的机制尚有待进一步研究。
3.6 微量元素
最近试验表明,在奶牛日粮中添加氨基酸螯合物代替无机微量元素可提高奶牛产奶量10.34%(P<0.05);提高乳蛋白含量3.4%(P<0.05);对乳脂率、非乳脂固形物含量和奶料比无影响。何瑞国等(2000)在奶牛日粮中添加复合微量元素缓释丸,试验结果表明,产奶量提高8.95%(P<0.01);乳糖、乳脂及乳蛋白分别提高8.12%(P<0.01)、11.01%(P<0.05)、5.79%(P<0.05)。日粮中补充铜、锌、硒三种微量元素,产奶量明显提高,乳蛋白率、乳脂率均有不同程度的增加(郭宏武等,1999)。也有资料报道,添加铜、锌、硒、碘复合微量元素有降低乳脂率的趋势,对乳蛋白的含量未见影响,而对乳糖含量稍微有增加的趋势(刘明祥等,1999)。而Campbell等(1999)试验表明,添加钴、铜、锰、锌不影响奶组成。铜可能会改变牛奶中脂肪酸的组成(Engle等,2001;Morales等,2000)。有关添加矿物质的作用效果和作用机理有待进一步研究。
4 小结
综上所述,影响牛奶产量和乳蛋白的营养和非营养因素很多,而关键营养因素就是奶牛产奶性能的不断提高对能量的需要增加,而奶牛日粮的能量由需要量和采食量决定,粗饲料品质差决定了不得不减少其用量,增加精料用量以满足能量需要。此外,可以通过日粮中添加过瘤胃脂肪来增加奶牛对能量的需要,但乳蛋白浓度有降低的可能,而乳脂率却提高,与过瘤胃蛋白或烟酸合用可缓解这一问题。日粮蛋白质的来源和氨基酸的组成对乳蛋白有很大的影响,但直接提高日粮的蛋白水平对提高乳蛋白效果不明显,因为日粮蛋白一般在瘤胃中被分解,因此必须使用过瘤胃蛋白质或保护性氨基酸才能改善瘤胃微生物的合成,提高小肠蛋白质的流量和氨基酸的组成,从而提高小肠对氨基酸的吸收和利用,满足乳腺组织蛋白质合成对氨基酸的需要,最终使乳蛋白合成达到最优化。但是,乳蛋白是遗传因子、日粮营养和管理及内分泌等综合调控的结果,尽管通过饲养和营养方法进行综合调控是一条简单易行、效果明显的途径,但不一定能取得理想的效果。在提高牛奶中蛋白质浓度的同时往往会降低牛奶中其它指标的含量,因而系统地研究这些因素并人为地根据具体情况进行综合调控,有效地提高乳蛋白的产量和含量,以满足人类对奶产品不断变化的需求显得尤为重要。
| 联系人: | 司达特 |
|---|---|
| 电话: | 400-698-2588/13910009918 |
| Email: | info@storti.com.cn |
| 地址: | 北京市朝阳区胜古中路企发大厦F座9层 |